Fosforilación Oxidativa
Acabas de leer sobre dos vías en el catabolismo de la glucosa-glicólisis y el ciclo del ácido cítrico-que generan ATP. Sin embargo, la mayor parte del ATP generado durante el catabolismo aeróbico de la glucosa no se genera directamente de estas vías. Más bien, se deriva de un proceso que comienza con el paso de electrones a través de una serie de reacciones químicas a un aceptor final de electrones, el oxígeno. Estas reacciones tienen lugar en complejos proteicos especializados situados en la membrana interna de las mitocondrias de los organismos eucariotas y en la parte interna de la membrana celular de los organismos procariotas. La energía de los electrones se recoge y se utiliza para generar un gradiente electroquímico a través de la membrana mitocondrial interna. La energía potencial de este gradiente se utiliza para generar ATP. La totalidad de este proceso se denomina fosforilación oxidativa.
La cadena de transporte de electrones (figura 4.19 a) es el último componente de la respiración aeróbica y es la única parte del metabolismo que utiliza el oxígeno atmosférico. El oxígeno se difunde continuamente en las plantas con este fin. En los animales, el oxígeno entra en el cuerpo a través del sistema respiratorio. El transporte de electrones es una serie de reacciones químicas que se asemeja a una brigada de cubos en la que los electrones pasan rápidamente de un componente al siguiente, hasta el punto final de la cadena donde el oxígeno es el aceptor final de electrones y se produce agua. Hay cuatro complejos compuestos por proteínas, etiquetados del I al IV en la Figura 4.19 c, y la agregación de estos cuatro complejos, junto con los portadores de electrones móviles y accesorios asociados, se denomina cadena de transporte de electrones. La cadena de transporte de electrones está presente en múltiples copias en la membrana mitocondrial interna de los eucariotas y en la membrana plasmática de los procariotas. En cada transferencia de un electrón a través de la cadena de transporte de electrones, el electrón pierde energía, pero con algunas transferencias, la energía se almacena como energía potencial utilizándola para bombear iones de hidrógeno a través de la membrana mitocondrial interna hacia el espacio intermembrana, creando un gradiente electroquímico.
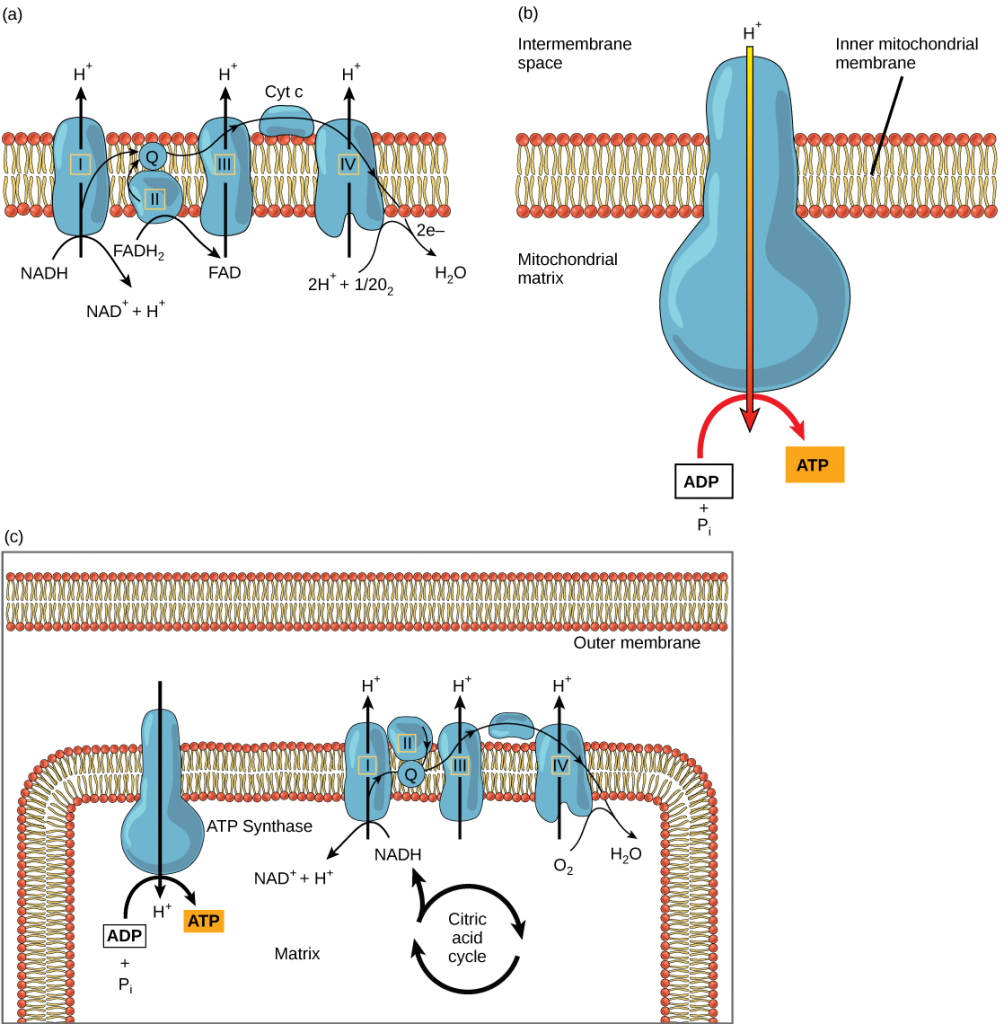
El cianuro inhibe la citocromo c oxidasa, un componente de la cadena de transporte de electrones. Si se produce un envenenamiento por cianuro, ¿esperarías que el pH del espacio intermembrana aumentara o disminuyera? ¿Qué efecto tendría el cianuro en la síntesis de ATP?
Los electrones del NADH y del FADH2 pasan a los complejos proteicos de la cadena de transporte de electrones. Al pasar de un complejo a otro (hay un total de cuatro), los electrones pierden energía, y parte de esa energía se utiliza para bombear iones de hidrógeno de la matriz mitocondrial al espacio intermembranal. En el cuarto complejo proteico, los electrones son aceptados por el oxígeno, el aceptor terminal. El oxígeno, con sus electrones adicionales, se combina entonces con dos iones de hidrógeno, aumentando aún más el gradiente electroquímico, para formar agua. Si no hubiera oxígeno en la mitocondria, los electrones no podrían ser eliminados del sistema, y toda la cadena de transporte de electrones retrocedería y se detendría. La mitocondria sería incapaz de generar nuevo ATP de esta manera, y la célula acabaría muriendo por falta de energía. Esta es la razón por la que debemos respirar para obtener nuevo oxígeno.
En la cadena de transporte de electrones, la energía libre de la serie de reacciones que acabamos de describir se utiliza para bombear iones de hidrógeno a través de la membrana. La distribución desigual de los iones H+ a través de la membrana establece un gradiente electroquímico, debido a la carga positiva de los iones H+ y a su mayor concentración en un lado de la membrana.
Los iones de hidrógeno se difunden a través de la membrana interna a través de una proteína integral de la membrana llamada ATP sintasa (figura 4.19 b). Esta compleja proteína actúa como un diminuto generador, que gira por la fuerza de los iones de hidrógeno que se difunden a través de ella, bajando su gradiente electroquímico desde el espacio intermembrana, donde hay muchos iones de hidrógeno que se repelen mutuamente, hasta la matriz, donde hay pocos. El giro de las piezas de esta máquina molecular regenera el ATP a partir del ADP. Este flujo de iones de hidrógeno a través de la membrana a través de la ATP sintasa se denomina quimiosmosis.
La quimiosmosis (figura 4.19 c) se utiliza para generar el 90 por ciento del ATP fabricado durante el catabolismo aeróbico de la glucosa. El resultado de las reacciones es la producción de ATP a partir de la energía de los electrones retirados de los átomos de hidrógeno. Estos átomos formaban originalmente parte de una molécula de glucosa. Al final del sistema de transporte de electrones, éstos se utilizan para reducir una molécula de oxígeno a iones de oxígeno. Los electrones adicionales de los iones de oxígeno atraen a los iones de hidrógeno (protones) del medio circundante y se forma agua. La cadena de transporte de electrones y la producción de ATP a través de la quimiosmosis se denominan colectivamente fosforilación oxidativa.