
Esta página começará com a bryophyte…
… ciclo de vida em resumo.
Um esporo de bryophyte é haplóide. Uma célula haplóide tem um conjunto de cromossomas, uma célula diplóide tem dois. Nos humanos, o óvulo e o espermatozóide são haplóides. Quando se unem, a célula resultante é diplóide e o embrião subsequente continua o seu desenvolvimento como indivíduo diplóide. Cada progenitor contribuiu com um conjunto de cromossomas.
Um esporo bryophyte germina e produz um tapete frequentemente parecido com algas, chamado protonema (protonemata plural) e a fase foliar ou tálóide do gametófito desenvolve-se a partir da fase protonemal. Os protonematos são quase sempre efémeros, mas há excepções. Entre os musgos há um pequeno número de espécies em que os protonematos são persistentes e as plantas folhosas são efémeras. O esporo foi haplóide, assim como o protonema resultante e as fases folhosas ou tálidas que se lhe seguiram. Os órgãos sexuais masculino e feminino, a anteridia e a arquegónia, respectivamente, são produzidos nas plantas gametófitas. Os espermatozóides haplóides são libertados da anteridia e quando um espermatozóide haplóide atinge um óvulo haplóide num arquegónio, o óvulo é fertilizado para produzir uma célula diplóide. Esta célula irá desenvolver-se para o esporófito diplóide. Num grande número de bryophytes, o tecido do esporófito embrionário diferencia-se. Tipicamente haverá um pé, que ancora o esporófito embrionário ao gametófito de suporte, e no final oposto ao pé é a parte que se desenvolverá na cápsula do esporófito. No meio há tipicamente a parte que se tornará o talo (ou seta) que suporta a cápsula do esporo, embora num certo número de bryophytes a seta seja curta ou mesmo inexistente. Quando a esporófita tiver amadurecido, a cápsula de esporos conterá esporos haplóides.
O diagrama seguinte dá um exemplo de um ciclo de vida de bryophyte, o do fungo Funaria hygrometrica.
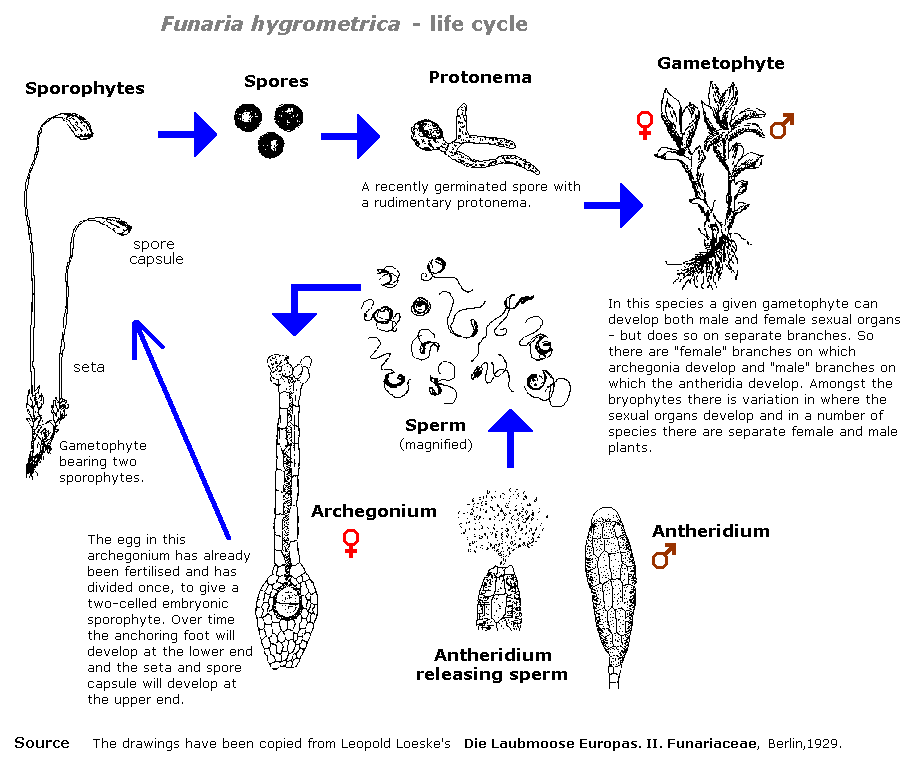 br>Imprima este diagrama como um PDF A4
br>Imprima este diagrama como um PDF A4
Os esporos produzidos numa cápsula de esporos são o resultado da REPRODUÇÃO SEXUAL. As briófitas são adeptas da REPRODUÇÃO VEGETATIVA e os propágulos vegetativos produzem frequentemente protonematos em vez de se desenvolverem directamente na fase gametófita foliar ou tálóide.
As briófitas são frequentemente ditas como mostrando uma alternância de gerações. Pode ver-se referências à “geração gametófita” e à “geração esporófita” alternando. A primeira coisa a notar é que a “geração” está a ser utilizada de uma forma bastante diferente da sua utilização quotidiana. Os pais (organismos diplóides) referem-se aos seus filhos (também organismos diplóides) como a “geração seguinte”. O segundo ponto digno de nota é que muitos gametófitos briófitos produzem prontamente descendência sem necessidade de esporófitos. O terceiro ponto a não ter em conta é que muitos organismos apresentam estados haplóides e diplóides. Nos humanos, o estado haplóide está confinado aos óvulos e espermatozóides e estes são incapazes de uma existência independente. Nas plantas em flor, os ovos e o pólen constituem o estado haplóide e também não são capazes de uma existência independente. Os seres humanos e as plantas floríferas são exemplos de organismos diplóides dominantes. Pelo contrário, nas criófitas, o estado haplóide é capaz de existência independente e é tipicamente muito mais duradouro do que o estado esporófito. As briófitas são exemplos de organismos haplóides-dominantes.
O resto desta página é dedicada principalmente ao tema do desenvolvimento do gametófito, desde os esporos até ao estádio de folhas ou talos. Esta página termina com algumas interessantes ligações web. Existe uma página separada de DESENVOLVIMENTO DE SPOROPHYTE.
Esporo a protonema
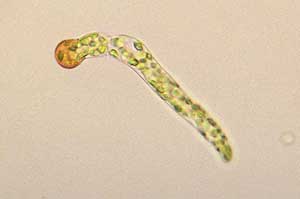 br> Esporo de Funaria germinativo
br> Esporo de Funaria germinativo p>Germinação de um esporo tipicamente produz um pequeno e filamentoso crescimento chamado tubo de germinação. A fotografia mostra um esporo germinativo de um musgo no género Funaria. O esporo é o objecto esférico acastanhado e um tubo de germinação cresceu a partir dele. O tubo mostrado nesta foto já tem várias células e também se podem ver muitos cloroplastos verdes, cheios de clorofila, nos quais se realiza a fotossíntese. O tubo inicial alonga-se e ramifica-se e esta foto![]() mostra os complexos filamentosos que se desenvolveram a partir de três esporos de Funaria. O crescimento precoce em muitos protonemas musgo consiste em tubos filamentosos com paredes incolores e com as paredes cruzadas entre as células vizinhas em ângulo recto na direcção do longo eixo do filamento. Esta fase é chamada cloronema. Mais tarde, desenvolve-se tipicamente um calulonema. O crescimento ainda é filamentoso mas agora as paredes são pigmentadas (tipicamente castanho-avermelhado) e as paredes cruzadas são obliquamente orientadas.
mostra os complexos filamentosos que se desenvolveram a partir de três esporos de Funaria. O crescimento precoce em muitos protonemas musgo consiste em tubos filamentosos com paredes incolores e com as paredes cruzadas entre as células vizinhas em ângulo recto na direcção do longo eixo do filamento. Esta fase é chamada cloronema. Mais tarde, desenvolve-se tipicamente um calulonema. O crescimento ainda é filamentoso mas agora as paredes são pigmentadas (tipicamente castanho-avermelhado) e as paredes cruzadas são obliquamente orientadas.
Os seguintes diagramas estilizados mostram algumas das variações.
No primeiro par de diagramas o esporo de musgo cinzento germinou e produziu um cloronema (mostrado como ![]() , com as linhas pretas indicando as paredes cruzadas entre as células vizinhas). Logo após a germinação pode ver-se ramos no cloronema e no diagrama à esquerda pode ver-se o desenvolvimento de um calunema, com as suas paredes cruzadas obliquamente orientadas, mostrado por
, com as linhas pretas indicando as paredes cruzadas entre as células vizinhas). Logo após a germinação pode ver-se ramos no cloronema e no diagrama à esquerda pode ver-se o desenvolvimento de um calunema, com as suas paredes cruzadas obliquamente orientadas, mostrado por ![]() . Também se pode ver o início do crescimento das folhas (
. Também se pode ver o início do crescimento das folhas ( ![]() ) a partir do calulonema. O cloronema é rico em cloroplastos mas o caulonema tem relativamente poucos. A fase foliar do gametófito desenvolve-se tipicamente a partir de um caulonema mas, como mostra o diagrama à direita, em alguns musgos desenvolve-se o crescimento foliar a partir do cloronema. É possível que um caulonema dê origem a cloronema fresco, como mostra um cloronema parcialmente pontilhado no diagrama à esquerda.
) a partir do calulonema. O cloronema é rico em cloroplastos mas o caulonema tem relativamente poucos. A fase foliar do gametófito desenvolve-se tipicamente a partir de um caulonema mas, como mostra o diagrama à direita, em alguns musgos desenvolve-se o crescimento foliar a partir do cloronema. É possível que um caulonema dê origem a cloronema fresco, como mostra um cloronema parcialmente pontilhado no diagrama à esquerda.

É importante que compreenda que todos os diagramas de protonema nesta página mostram simplesmente as várias fases iniciais do crescimento do gametófito e as suas relações umas com as outras. Não os tome como representações realistas do desenvolvimento real do gametófito. Por exemplo, tomados à letra, os dois diagramas imediatamente acima mostram os botões folhosos que se desenvolvem sobre um pequeno protonema, aparentemente muito pouco tempo após a germinação dos esporos. De facto, um protonema de musgo pode ser muito extenso antes dos botões folhosos começarem a desenvolver-se.
 Na maioria dos musgos o protonema é filamentoso mas em alguns géneros é tálóide. O diagrama (à direita) mostra um exemplo do género Sphagnum. Há um cloronema curto e filamentoso, mas logo se desenvolve para uma forma de talo. A área da talose, embora composta por muitas células, é apenas uma célula de espessura. Os crescimentos azuis são rizóides, com paredes transversais oblíquas. Note-se que existe uma variação no género. Em algumas espécies de Sphagnum, o protonema da talose é em forma de fita e noutras é altamente lobulado. Pode haver protonematos secundários de talose que se desenvolvem no final de um pequeno filamento do protonema inicial.
Na maioria dos musgos o protonema é filamentoso mas em alguns géneros é tálóide. O diagrama (à direita) mostra um exemplo do género Sphagnum. Há um cloronema curto e filamentoso, mas logo se desenvolve para uma forma de talo. A área da talose, embora composta por muitas células, é apenas uma célula de espessura. Os crescimentos azuis são rizóides, com paredes transversais oblíquas. Note-se que existe uma variação no género. Em algumas espécies de Sphagnum, o protonema da talose é em forma de fita e noutras é altamente lobulado. Pode haver protonematos secundários de talose que se desenvolvem no final de um pequeno filamento do protonema inicial.
 O tipo de desenvolvimento Hedwigia é mostrado pela espécie Hedwigia ciliata. Os esporos deste musgo têm paredes espessas e o esporo divide-se dentro da parede do esporo, levando à existência de duas células dentro da parede do esporo esticado. O crescimento sedutor leva à ruptura da parede do esporo esticado e ao desenvolvimento de um componente protonemal constituído por entre 20 e 25 células globosas. A isto chama-se um protonema maciço e esta forma de desenvolvimento é mostrada no diagrama seguinte (à direita), com alguns fragmentos de parede de esporos cinzentos escuros visíveis no protonema maciço. Não há cloronema e o calulonema desenvolve-se a partir do protonema maciço. Os botões de gametófitos folhosos podem desenvolver-se em qualquer um ou em ambos os protonemas e caulonemas maciços.
O tipo de desenvolvimento Hedwigia é mostrado pela espécie Hedwigia ciliata. Os esporos deste musgo têm paredes espessas e o esporo divide-se dentro da parede do esporo, levando à existência de duas células dentro da parede do esporo esticado. O crescimento sedutor leva à ruptura da parede do esporo esticado e ao desenvolvimento de um componente protonemal constituído por entre 20 e 25 células globosas. A isto chama-se um protonema maciço e esta forma de desenvolvimento é mostrada no diagrama seguinte (à direita), com alguns fragmentos de parede de esporos cinzentos escuros visíveis no protonema maciço. Não há cloronema e o calulonema desenvolve-se a partir do protonema maciço. Os botões de gametófitos folhosos podem desenvolver-se em qualquer um ou em ambos os protonemas e caulonemas maciços.
O tipo de desenvolvimento Drummondia é semelhante ao tipo Hedwigia, excepto que o protonema maciço, de 8-20 células, ocorre dentro de uma parede de esporos esticada mas não rompida e não há caulonema. Uma vez que o desenvolvimento ocorre dentro da parede dos esporos, chama-se germinação endosporosa. Pelo contrário, os outros exemplos dados acima mostram uma germinação exosporosa, uma vez que todo o desenvolvimento protonémico tem lugar fora do esporo.
O desenvolvimento exosporoso e endosporoso ocorre também nos fígados e nos chifres, mas o calulonema só se encontra nos musgos. Nos musgos e nos chifres a germinação exosporosa é muito mais comum que a exosporosa, mas nos hepáticos a germinação endosporosa é mais comum. Os protonematos de fígado mostram uma variedade de formas protonematosas (filamentosos, em forma de fita, discóides, maciços) mas os protonematos de fígado são geralmente insignificantes em extensão, tal como os de chifres![]() .
.
Mais crescimento de gametófitos
Crescimento de gametófitos bryophytos está sob controlo de células apicais. Estas são células que se dividem repetidamente para formar novas células. Uma célula apical tem uma face frontal e até quatro faces posteriores. As faces posteriores estão ligadas ao resto da gametófita, mas a face dianteira é livre. As novas células formam-se apenas ao longo das faces posteriores. As células recém-formadas são pequenas para começar, mas depois expandem-se para que o gametófito se estenda na direcção da face anterior da célula apical. As células apicais são fundamentais para o crescimento do gametófito. Cada caule, folha, ramo do talo, etc., tem a sua célula ou células apicais. Numa bryophyte folhosa as folhas crescem fora dos caules. Na ponta de crescimento de cada caule, existe uma célula apical. Há também uma célula apical no ápice de cada folha em crescimento. Nas bryófitas de talo pode haver várias células apicais, lado a lado, numa fila curta na margem do talo. O ESTUDO DE CASO APICAL CÉLULAS entra em mais detalhe.
Gametophytes produzem o esperma masculino e os óvulos femininos. Os espermatozóides são produzidos dentro de anterídios e os ovos são produzidos em arquegónias. Dependendo da espécie em questão, tanto a anteridia como a arquegónia podem ser produzidas no mesmo gametófito ou em gametófitos separados. No primeiro caso, com ambos os tipos de órgãos sexuais num gametófito, o gametófito é bissexual (ou monóico). No segundo caso, existem plantas masculinas e femininas separadas, pelo que cada gametófito é unissexual (ou dióico). Há muitas espécies dióicas e muitas espécies monóicas. Dentro das bryophytes monóicas as anterídios e arquegónias podem estar no mesmo ramo ou em ramos diferentes ou lóbulos de talo, mais uma vez dependendo da espécie. Pessoas com algum conhecimento botânico ou zoológico podem ter-se deparado com os mesmos termos dioeciosos e monóicos. Encontrará um relato da história e dos usos variados dos quatro termos no ESTUDO DO CASO -OICY. Nesse link encontrará também descrições e termos para os vários arranjos dos órgãos sexuais das espécies monóicas. Essa discussão é aí colocada para os interessados em mais detalhes, mas não é necessária para uma compreensão básica das bryophytes. O mais importante é simplesmente estar ciente de que existem espécies unissexuais e bissexuais.
 br>Gigaspermum repens
br>Gigaspermum repens p> nas espécies dióicas os gametófitos macho e fêmea podem ter muito o mesmo tamanho ou com machos mais pequenos, por vezes de forma dramática. Nesta foto da espécie de musgo Gigaspermum repens pode facilmente ver as plantas fêmeas de folhas largas com as suas cápsulas de esporos. As plantas machos são as plantas muito mais pequenas que são mais facilmente vistas em direcção às margens da fotografia. Esta não é de forma alguma a disparidade de tamanho mais notável, pois existem espécies de musgo em que as plantas machos não são mais do que pequenos apêndices nas folhas das plantas fêmeas.
Alguns links interessantes
O website do Departamento de Biologia de Organismos e Sistemas da Universidade de Oviedo (nas Astúrias, Espanha) tem representações belamente desenhadas dos ciclos de vida de três bryophytes. As legendas estão em espanhol, mas devem ser relativamente facilmente compreendidas por pessoas sem conhecimentos de espanhol. Muitos dos termos técnicos são semelhantes em outras línguas e haveria poucas palavras que seria necessário procurar num dicionário bilingue. Aqui estão links específicos para os três diagramas:
Phaeoceros laevis, um hornwort.
Marchantia polymorpha, uma talocha complexa.
Polytrichum, um musgo.